This is chapter 1 of the "The
Phenomenon of Science" by Valentin F.
Turchin
Contents:
IN THE PROCESS of the evolution of life, as far as we know, the total mass of living matter has always been and is now increasing and growing more complex in its organization. To increase the complexity of the organization of biological forms, nature operates by trial and error. Existing forms are reproduced in many copies, but these are not identical to the original. Instead they differ from it by the presence of small random variations. These copies then serve as the material for natural selection. They may act as individual living beings, in which case selection leads to the consolidation of useful variations, or elements of more complex forms, in which case selection is also directed to the structure of the new form (for example, with the appearance of multicellular organisms). In both cases selection is the result of the struggle for existence, in which more viable forms supplant less viable ones.
This mechanism of the development of life, which was discovered by Charles Darwin, may be called the basic law of evolution. It is not among our purposes to substantiate or discuss this law from the point of view of those laws of nature which could be declared more fundamental. We shall take the basic law of evolution as given.
THE HISTORY OF LIFE before the appearance of the human being can be broken into two periods, which we shall call the "chemical'' era and the ''cybernetic'' era. The bridge between them is the emergence of animals with distinct nervous systems, including sense organs, nerve fibers for transmitting information, and nerve centers (nodes) for converting this information. Of course, these two terms do not signify that the concepts and methods of cybernetics are inapplicable to life in the ''chemical" era; it is simply that the animal of the ''cybernetic'' era is the classical object of cybernetics, the one to which its appearance and establishment as a scientific discipline are tied.
We shall review the history and logic of evolution in the precybernetic period only in passing, making reference to the viewpoints of present-day biologists.[1] Three stages can be identified in this period.
In the first stage the chemical foundations of life are laid. Macromolecules of nucleic acids and proteins form with the property of replication, making copies or ''prints'' where one macromolecule serves as a matrix for synethesizing a similar macromolecule from elementary radicals. The basic law of evolution, which comes into play at this stage, causes matrices which have greater reproductive intensity to gain an advantage over matrices with lesser reproductive intensity, and as a result more complex and active macromolecules and systems of macromolecules form. Biosynthesis demands free energy. Its primary source is solar radiation. The products of the partial decay of life forms that make direct use of solar energy (photosynthesis) also contain a certain reserve of free energy which may be used by the already available chemistry of the macromolecule. Therefore, this reserve is used by special forms for which the products of decay serve as a secondary source of free energy. Thus the division of life into the plant and animal worlds arises.
The second stage of evolution is the appearance and development of the motor apparatus in animals.
Plants and animals differ fundamentally in the way they obtain energy. With a given level of illumination the intensity of absorption of solar energy depends entirely on the amount of plant surface, not on whether it moves or remains stationary. Plants were refined by the creation of outlying light catchers--green leaves secured to a system of supports and couplings (stems, branches, and the like). This design works very well, ensuring a slow shift in the green surfaces toward the light which matches the slow chance in illumination.
The situation is entirely different with animals, in particular with the most primitive types such as the amoeba. The source of energy-- food--fills the environment around it. The intake of energy is determined by the speed at which food molecules are diffused through the shell that separates the digestive apparatus from the external environment. The speed of diffusion depends less on the size of the surface of the digestive apparatus than on the movement of this surface relative to the environment; therefore it is possible for the animal to take in food from different sectors of the environment. Consequently, even simple, chaotic movement in the environment or, on the other hand, movement of the environment relative to the organism (as is done, for example, by sponges which force water through themselves by means of their cilia) is very important for the primitive animal and, consequently, appears in the process of evolution. Special forms emerge (intracellular formations in one-celled organisms and ones containing groups of cells in multicellular organisms) whose basic function is to produce movement.
In the third stage of evolution the movements of animals become directed and the incipient forms of sense organs and nervous systems appear in them. This is also a natural consequence of the basic law. It is more advantageous for the animal to move in the direction where more food is concentrated, and in order for it to do so it must have sensors that describe the state of the external environment in all directions (sense organs) and information channels for communication between these sensors and the motor apparatus (nervous system). At first the nervous system is extremely primitive. Sense organs merely distinguish a few situations to which the animal must respond differently. The volume of information transmitted by the nervous system is slight and there is no special apparatus for processing the information. During the process of evolution the sense organs become more complex and deliver an increasing amount of information about the external environment. At the same time the motor apparatus is refined, which makes ever-increasing demands on the carrying capacity of the nervous system. Special formations appear--nerve centers which convert information received from the sense organs into information controlling the organs of movement. A new era begins: the ''cybernetic'' era.
TO ANALYZE evolution in the cybernetic period and to discover the laws governing the organization of living beings in this period (for brevity we will call them "cybernetic animals'') we must introduce certain fundamental concepts and laws from cybernetics.
The term ''cybernetics'' itself was, of course, introduced by Norbert Wiener, who defined it descriptively as the theory of relationships and control in the living organism and the machine. As is true in any scientific discipline, a more precise definition of cybernetics requires the introduction of its basic concepts. Properly speaking, to introduce the basic concepts is the same as defining a particular science, for all that remains to be added is that a description of the world by means of this system of concepts is, in fact, the particular, concrete science.
Cybernetics is based above all on the concept of the system, a certain material object which consists of other objects which are called subsystems of the given system. The subsystem of a certain system may, in its turn, be viewed as a system consisting of other subsystems. To be precise, therefore, the meaning of the concept we have introduced does not lie in the term ''system'' by itself, that is, not in ascribing the property of ''being a system'' to a certain object (this is quite meaningless, for any object may be considered a system), but rather in the connection between the terms ''system'' and "subsystem," which reflects a definite relationship among objects.
The second crucial concept of cybernetics is the concept of the state of a system (or subsystem). Just as the concept of the system relies directly on our spatial intuition, the concept of state relies directly on our intuition of time and it cannot be defined except by referring to experience. When we say that an object has changed in some respect we are saying that it has passed into a different state. Like the concept of system. the concept of state is a concealed relationship: the relationship between two moments in time. If the world were immobile the concept of state would not occur, and in those disciplines where the world is viewed statically, for example in geometry, there is no concept of state.
Cybernetics studies the organization of systems in space and time, that is, it studies how subsystems are connected into a system and how change in the state of some subsystems influences the state of other subsystems. The primary emphasis, of course, is on organization in time which, when it is purposeful, is called control. Causal relations between states of a system and the characteristics of its behavior in time which follow from this are often called the dynamics of the system, borrowing a term from physics. This term is not applicable to cybernetics. because when we speak of the dynamics of a system we are inclined to view it as something whole, whereas cybernetics is concerned mainly with investigating the mutual influences of subsystems making up the particular system. Therefore, we prefer to speak of organization in time, using the term dynamic description only when it must be juxtaposed to the static description which considers nothing but spatial relationships among subsystems.
A cybernetic description may have different levels of detail. The same system may be described in general outline, in which it is broken down into a few large subsystems or "blocks,'' or in greater detail, in which the structure and internal connections of each block are described. But there is always some final level beyond which the cybernetic description does not apply. The subsystems of this level are viewed as elementary and incapable of being broken down into constituent parts. The real physical nature of the elementary subsystems is of no interest to the cyberneticist, who is concerned only with how they are interconnected. The nature of two physical objects may be radically different, but if at some level of cybernetic description they are organized from subsystems in the same way (considering the dynamic aspect!), then from the point of view of cybernetics they can be considered, at the given level of description. identical. Therefore, the same cybernetic considerations can be applied to such different objects as a radar circuit, a computer program, or the human nervous system.
THE STATE OF A SYSTEM is defined through the aggregate of states of all its subsystems, which in the last analysis means the elementary subsystems. There are two types of elementary subsystems: those with a finite number of possible states, also called subsystems with discrete states, and those with an infinite number, also called subsystems with continuous states. The wheel of a mechanical calculator or taxi meter is an example of a subsystem with discrete states. This wheel is normally in one of 10 positions which correspond to the 10 digits between 0 and 9. From time to time it turns and passes from one state into another. This process of turning does not interest us. The correct functioning of the system (of the calculator or meter) depends entirely on how the ''normal'' positions of the wheels are interconnected, while how the change from one position (state) to another takes place is inconsequential. Therefore we can consider the calculator as a system whose elementary subsystems can only be in discrete states. A modern high-speed digital computer also consists of subsystems (trigger circuits) with discrete states. Everything that we know at the present time regarding the nervous systems of humans and animals indicates that the interaction of subsystems (neurons) with discrete states is decisive in their functioning.
On the other hand, a person riding a bicycle and an anal computer are both examples of systems consisting of subsystems with continuous states. In the case of the bicycle rider these subsystems are all the parts of the bicycle and human body which are moving relative to one another: the wheels, pedals, handlebar, legs, arms, and so on. Their states are their positions in space. These positions are described by coordinates (numbers) which can assume continuous sets of values.
If a system consists exclusively of subsystems with discrete states then the system as a whole must be a system with discrete states. We shall simply call such systems ''discrete systems,'' and we shall call systems with continuous sets of states ''continuous systems.'' In many respects discrete systems are simpler to analyze than continuous ones. Counting the number of possible states of a system, which plays an important part in cybernetics, requires only a knowledge of elementary arithmetic in the case of discrete systems. Suppose discrete system A consists of two subsystems a1 and a2; subsystem al may havea1 possible states, while subsystem a2 may have n2. Assuming that each state of system al can combine with each state of system a2 we find that N, the number of possible states of system A, is n1n2. If system A consists of m subsystems a1 where i = 1, 2, . . ., m, then
N = a2a2, . . . nm
From this point on we shall consider only discrete systems. In addition to the pragmatic consideration that they are simpler in principle than continuous systems, there are two other arguments for such a restriction.
First, all continuous systems can in principle be viewed as discrete systems with an extremely large number of states. In light of the knowledge quantum physics has given us, this approach can even be considered theoretically more correct. The reason why continuous systems do not simply disappear from cybernetics is the existence of a very highly refined apparatus for consideration of such systems: mathematical analysis, above all, differential equations.
Second, the most complex cybernetic systems, both those which have arisen naturally and those created by human hands, have invariably proved to be discrete. This is seen especially clearly in the example of animals. The relatively simple biochemical mechanisms that regulate body temperature, the content of various substances in the blood, and similar characteristics are continuous, but the nervous system is constructed according to the discrete principle.
WHY DO DISCRETE SYSTEMS prove to be preferable to continuous ones when it is necessary to perform complex functions? Because they have a much higher reliability. In a cybernetic device based on the principle of discrete states each elementary subsystem may be in only a small number of possible states, and therefore the system ordinarily ignores small deviations from the norm of various physical parameters of the system, reestablishing one of its permissible states in its ''primeval purity.'' In a continuous system, however, small disturbances continuously accumulate and if the system is too complex it ceases functioning correctly. Of course, in the discrete system too there is always the possibility of a breakdown, because small changes in physical parameters do lead to a finite probability that the system will switch to an ''incorrect'' state. Nonetheless, discrete systems definitely have the advantage. Let us demonstrate this with the following simple example.
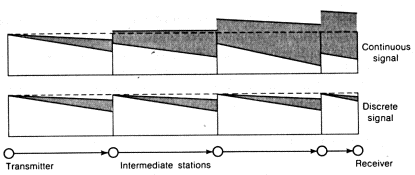
Suppose we must transmit a message by means of electric wire over a distance of, say, 100 kilometers (62 miles). Suppose also that we are able to set up an automatic station for every kilometer of wire and that this station will amplify the signal to the power it had at the previous station and, if necessary, convert the signal.
Figure 1.1. Transmission of a signal in continuous and discrete systems (The shaded part shows the area of signal ambiguity.)
We assume that the maximum signal our equipment permits us to send has a magnitude of one volt and that the average distortion of the signal during transmission from station to station (noise) is equal to 0.1 volt.
First let us consider the continuous method of data transmission. The content of the message will be the amount of voltage applied to the wire at its beginning. Owing to noise, the voltage at the other end of the wire--the message received--will differ from the initial voltage. How great will this difference be? Considering noise in different segments of the line to be independent, we find that after the signal passes the 100 stations the root-mean square magnitude of noise will be one volt (the mean squares of noise are summed). Thus, average noise is equal to the maximum signal, and it is therefore plain that we shall not in fact receive any useful information. Only by accident can the value of the voltage received coincide with the value of the voltage transmitted. For example, if a precision of 0.1 volt satisfies us the probability of such a coincidence is approximately 1/10.
Now let us look at the discrete variant. We shall define two "meaningful'' states of the initial segment of the wire: when the voltage applied is equal to zero and when it is maximal (one volt). At the intermediate stations we install automatic devices which transmit zero voltage on if the voltage received is less than 0.5 volt and transmit a normal one-volt signal if the voltage received is more than 0.5 volt. In this case, therefore, for one occasion (one signal) information of the "yes/no" type is transmitted (in cybernetics this volume of information is called one "bit" ). The probability of receiving incorrect information depends strongly on the law of probability distribution for the magnitude of noise. Noise ordinarily follows the so-called normal law. Assuming this law we can find that the probability of error in transmission from one station to the next (which is equal to the probability that noise will exceed 0.5 volt) is 0.25 . 10-6. Thus the probability of an error in transmission over the full length of the line is 0.25 . 10-4. To transmit the same message as was transmitted in the previous case--that is, a value between 0 and 1 with a precision of 0.1 of a certain quantity lying between 0 and l--all we have to do is send four ''yes/no'' type signals. The probability that there will be error in at least one of the signals is 0-4. Thus, with the discrete method the total probability of error is 0.01 percent, as against 90 percent for the continuous method.
WHEN WE BEGAN describing a concrete cybernetic system it was impossible not to use the term information--a word familiar and understandable in its informal conversational meaning. The cybernetic concept of information, however., has an exact quantitative meaning.
Let us imagine two subsystems A and B
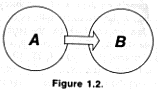
The two subsystems are interconnected in such a way that a change in the state of A leads to a change in the state of B. This can also be expressed as follows: A influences B. Let us consider the state of B at a certain moment in time t1 and at a later moment t2. We shall signify the first state as S1 and the second as S2. State S2 depends on state S1. The relationship of S2 to S1 is probabilistic, however, not unique. This is because we are not considering an idealized theoretical system governed by a deterministic law of movement but rather a real system whose states Si are the results of experimental data. With such an approach we may also speak of the law of movement, understanding it in the probabilistic sense--that is, as the conditional probability of state S2 at moment t2 on the condition that the system was in state S1 at moment t1. Now let us momentarily ignore the law of movement. We shall use N to designate the total number of possible states of subsystem B and imagine that conditions are such that at any moment in time system B can assume any of N states with equal probability, regardless of its state at the preceding moment. Let us attempt to give a quantitative expression to the degree (or strength) of the cause-effect influence of system A on such an inertialess and ''lawless'' subsystem B.
Suppose B acted upon by A switches to a certain completely determinate state. It is clear that the ''strength of influence'' which is required from A for this depends on N, and will be larger as N is larger. For example, if N= 2 then B, even if it is completely unrelated to A, when acted upon by random factors can switch with a probability of .5 to the very state A ''recommends.'' But if N = 109, when we have noticed such a coincidence we shall hardly doubt the influence of A on B. Therefore, some monotonic increasing function of N should serve as the measure of the 'strength of the influence'' of A on B. What this essentially means is that it serves as a measure of the intensity of the cause-effect relationship between two events, the state of A in the time interval from t1 to t2 and the state of B at t2. In cybernetics this measure is called the quantity of information transmitted from A to B between moments in time t1 and t2, and a logarithm serves as the monotonic increasing function. So, in our example, the quantity of information I passed from A to B is equal to log N.
Selection of the logarithmic function is determined by its property according to which
log N1N2 = log N1 + log N2
Suppose system A influences system B which consists of two independent subsystems B1 and B2with number of possible states N1 and N2 respectively.
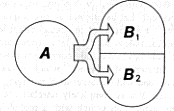
Then the number of states of system B is N1N2 and the quantity of information I that must be transmitted to system B in order for it to assume one definite state is, owing to the above-indicated property of the logarithm, the sum
I = log N1N2 = log N1 + log N2 = I1 + I2
where I1 and I2 are the quantities of information required by subsystems B1 + B2. Thanks to this property the information assumes definite characteristics of a substance; it spreads over the independent subsystems like a fluid filling a number of vessels. We are speaking of the joining and separation of information flows, information capacity, and information processing and storage.
The question of information storage is related to the question of the law of movement. Above we mentally set aside the law of movement in order to define the concept of information transmission. If we now consider the law of movement from this new point of view, it can be reduced to the transmission of information from system B at moment t1 to the same system B at moment t1. If the state of the system does not change with the passage of time, this is information storage. If state S2 is uniquely determined by S1 at a preceding moment in time the system is called fully deterministic. If S1 is uniquely determined by S2 the system is called reversible; for a reversible system it is possible in principle to compute all preceding states on the basis of a given state because information loss does not occur. If the system is not reversible information is lost. The law of movement is essentially something which regulates the flow of information in time from the system and back to itself.
Figure 1.4 shows the chart of information transmission from system A to system C through system B.
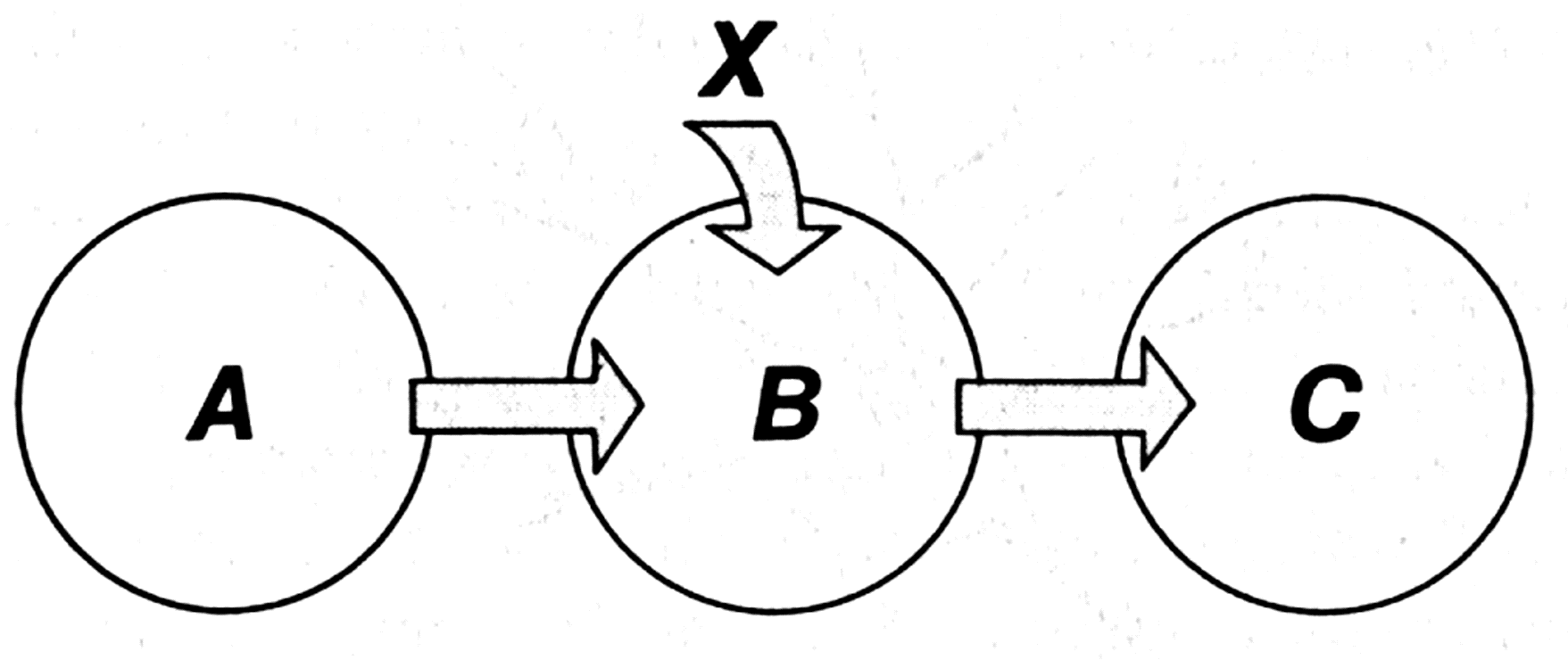
B is called the communication channel. The state of B can be influenced not only by the state of system A, but also by a certain uncontrolled factor X, which is called noise. The final state of system C in this case depends not only on the state of A, but also on factor X (information distortion). One more important diagram of information exchange is shown in figure 1.5.
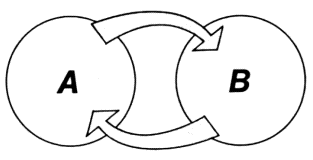
This is the so-called feedback diagram. The state of system A at t1 influences the state of B at t2, then the latter influences the state of A at t3. The circle of information movement is completed.
With this we conclude for now our familiarization with the general concepts of cybernetics and turn to the evolution of life on earth.
THE EXTERNAL APPEARANCE of a nerve cell (neuron) is shown schematically in figure 1.6.
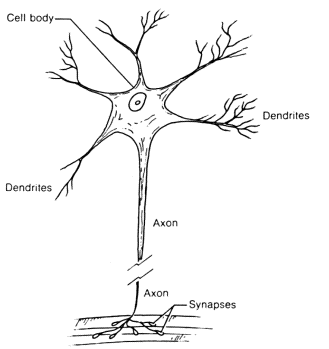
Figure 1.6. Diagram of the structure of a neuron.
A neuron consists of a fairly large (up to 0.1 mm) cell body from which several processes called dendrites spread, giving rise to finer and finer processes like the branching of a tree. In addition to the dendrites one other process branches out from the body of the nerve cell. This is the axon, which resembles a long, thin wire. Axons can be very long, up to a meter, and they end in treelike branching systems as do the dendrites. At the ends of the branches coming from the axon one can see small plates or bulblets. The bulblets of one neuron approach close to different segments of the body or dendrites of another neuron, almost touching them.
These contacts are called synapses and it is through them that neurons interact with one another. The number of bulblets approaching the dendrites of the single neuron may run into the dozens and even hundreds. In this way the neurons are closely interconnected and form a nerve net.
When one considers certain physicochemical properties (above all the propagation of electrical potential over the surface of the cell) one discovers the neurons can be in one of two states--the state of dormancy or the state of stimulation. From time to time, influenced by other neurons or outside factors, the neuron switches from one state to the other. This process takes a certain time, of course, so that an investigator who is studying the dynamics of the electrical state of a neuron, for example, considers it a system with continuous states. But the information we now have indicates that what is essential for the functioning of the nervous system as a whole is not the nature of switching processes but the very tact that the particular neurones are in one of these two states. Therefore, we may consider that the nerve net is a discrete system which consists of elementary subsystems (the neurons) with two states.
When the neuron is stimulated, a wave of electrical potential runs along the axon and reaches the bulblets in its branched tips. From the bulblets the stimulation is passed across the synapses to the corresponding sectors of the cell surface of other neurons. The behavior of a neuron depends on the state of its synapses. The simplest model of the functioning of the nerve net begins with the assumption that the state of the neuron at each moment in time is a single-valued function of the state of its synapses. It has been established experimentally that the stimulation of some synapses promotes stimulation of the cell, whereas the stimulation of other synapses prevents stimulation of the cell. Finally. certain synapses are completely unable to conduct stimulation from the bulblets and therefore do not influence the state of the neuron. It has also been established that the conductivity of a synapse increases after the first passage of a stimulus through it. Essentially a closing of the contact occurs. This explains how the system of communication among neurones, and consequently the nature of the nerve net's functioning, can change without a change in the relative positions of the neurons.
The idea of the neuron as an instantaneous processor of information received from the synapses is, of course, very simplified. Like any cell the neuron is a complex machine whose functioning has not yet been well understood. This machine has a large internal memory, and therefore its reactions to external stimuli may show great variety. To understand the general rules of the working of the nervous system, however. we can abstract from these complexities (and really, we have no other way to go!) and begin with the simple model outlined above.
A GENERALIZED DIAGRAM of the nerve system of the "cybernetic animal'' in its interaction with the environment is shown in figure 1.7.
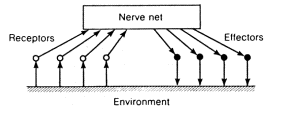
Figure 1.7. Nervous system of the "cybernetic animal"
Those sensory nerve cells which are stimulated by the action of outside factors are called receptors (that is, receivers) because they are the first to receive information about the state of the environment. This information enters the nerve net and is converted by it. As a result certain nerve cells called effectors are stimulated. Branches of the effector cells penetrate those tissues of the organism which the nervous system affects directly. Stimulation of the effector causes a contraction of the corresponding muscle or the stimulation of the activity of the appropriate gland. We shall call the state of all receptors at a certain moment in time the situation at that moment. (It would be more precise--if more cumbersome--to say the ''result of the effect of the situation on the sense organs.'') We will call the state of all the effectors the ''action.''. Therefore, the role of the nerve net is to convert a situation into an action.
It is convenient to take the term ''environment'' from figure 1.7 to mean not just the objects which surround the animal, but also its bone and muscle system and generally everything that is not part of the nervous system. This makes it unnecessary to give separate representations in the diagram to the animal body and what is not the body, especially because this distinction is not important in principle for the activity of the nervous system. The only thing that is important is that stimulation of the effectors leads to certain changes in the "environment.'' With this general approach to the problem as the basis of our consideration, we need only classify these changes as ''useful'' or ''harmful'' for the animal without going into further detail.
The objective of the nervous system is to promote the survival and reproduction of the animal. The nervous system works well when stimulation of the effectors leads to changes in the state of the environment that help the animal survive or reproduce, and it works badly when it leads to the reverse. With its increasing refinement in the process of evolution. the nervous system has performed this task increasingly well. How does it succeed in this? What laws does this process of refinement follow?
We will try to answer these questions by identifying in the evolution of the animal nervous system several stages that are clearly distinct from a cybernetic point of view and by showing that the transition from each preceding stage to each subsequent stage follows inevitably from the basic law of evolution. Because the evolution of living beings in the cybernetic era primarily concerns the evolution of their nervous systems, a periodization of the development of the nervous system yields a periodization of the development of life as a whole .
THE SIMPLEST VARIANT of the nerve net is when there is no net at all. In this case the receptors are directly connected to the effectors and stimulation from one or several receptors is transmitted to one or several effectors. We shall call such a direct connection between stimulation of a receptor and an etfector the simple reflex.
This stage, the third in our all-inclusive enumeration of the stages of evolution, is the bridge between the chemical and cybernetic eras. The Coelenterata are animals fixed at the level of the simple reflex. As an example let us take the hydra, which is studied in school as a typical representative of the Coelenterata. The body of a hydra has the shape of an elongated sac. Its interior, the coelenteron, is connected to the environment through a mouth, which is surrounded by several tentacles. The walls of the sac consist of two layers of cells: the inner layer (entoderm) and the outer layer (ectoderm). Both the ectoderm and the entoderm have many muscle cells which contain small fibers that are able to contract. thus setting the body of the hydra in motion. In addition, there are nerve cells in the ectoderm; the cells located closest to the surface are receptors and the cells which are set deeper, among. the muscles, are effectors. If a hydra is pricked with a needle it squeezes itself into a tiny ball. This is a simple reflex caused by transmission of the stimulation from the receptors to the effectors.
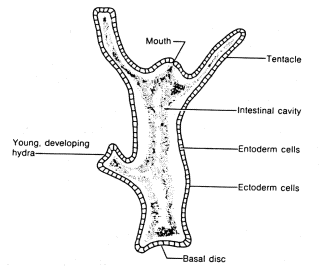
Figure 1.8. The structure of the hydra.
But the hydra is also capable of much more complex behavior. After it has captured prey, the hydra uses its tentacles to draw the prey to its mouth and then swallows the prey. This behavior can also be explained by the aggregate action of simple reflexes connecting effectors and receptors locally. within small segments of the body. For example, the following model of a tentacle explains its ability to wrap itself around captured objects.
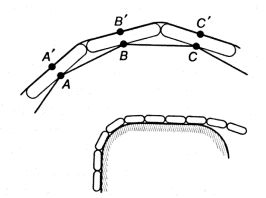
Figure 1.9. Model of a tentacle
Let us picture a certain number of links connected by hinges (for simplicity we shall consider a two-dimensional picture). Points A and B, A ' and B', B and C, and B' and C', etc. are interconnected by strands which can contract (muscles). All these points are sensitive and become stimulated when they touch an object (receptors). The stimulation of each point causes a contraction of the two strands connected to it (reflex).
THE SIMPLE REFLEX relationship between the stimulated cell and the muscle cell arises naturally, by the trial and error method, in the process of evolution. If the correlation between stimulation of one cell and contraction of another proves useful for the animal, then this correlation becomes established and reinforced. Where interconnected cells are mechanically copied in the process of growth and reproduction, nature receives a system of parallel-acting simple reflexes resembling the tentacle of the hydra. But when nature has available a large number of receptors and effectors which are interconnected by pairs or locally. there is a " temptation" to make the system of connections more complex by introducing intermediate neurons. This is advantageous because where there is a system of connections among all neurons, forms of behavior that are not possible where all connections are limited to pairs or localities now become so. This point can be demonstrated by a simple calculation of all the possible methods of converting a situation into an action with each method of interconnection. For example, assume that we have n receptors and effectors connected by pairs. In each pair the connection may be positive (stimulation causes stimulation and dormancy evokes dormancy) or negative (stimulation evokes dormancy and dormancy causes stimulation). In all, therefore, 2n variants are possible, which means 2n variants of behavior. But if we assume that the system of connections can be of any kind, which is to say that the state of each effector (stimulation or dormancy) can depend in any fashion on the state of all the receptors, then a calculation of all possible variants of behavior yields the number *2(2^n)n, which is immeasurably larger than 2n .
Exactly the same calculation leads to the conclusion that joining any subsystems which join independent groups of receptors and effectors into a single system always leads to an enormous increase in the number of possible variants of behavior. Throughout the entire course of the history of life, therefore, the evolution of the nervous system has progressed under the banner of increasing centralization.
But 'centralization'' can mean different thins. If all neurons are connected in one senselessly confused clump, then the system-- despite its extremely ''centralized'' nature--will hardly have a chance to survive in the struggle for existence. Centralization poses the following problem: how to select from all the conceivable ways of joining many receptors with many effectors (by means of intermediate neurons if necessary) that way which will correlate a correct action (that is, one useful for survival and reproduction) to each situation? After all, a large majority of the ways of interconnection do not have this characteristic.
We know that nature takes every new step toward greater complexity in living structures by the trial and error method. Let us see what direct application of the trial and error method to our problem yields. As an example we shall consider a small system consisting of 100 receptors and 100 effectors. We shall assume that we have available as many neurons as needed to create an intermediate nerve net and that we are able to determine easily whether the particular method of connecting neurons produces a correct reaction to each situation. We shall go through all conceivable ways of connection until we find the one we need. Where n = 100 the number of functionally different nerve nets among n receptors and n effectors is
2(2^n)n =~ 10(10^32)
This is an inconceivably large number. We cannot sort through such a number of variants and neither can Mother Nature. If every atom in the entire visible universe were engaged in examining variants and sorting them at a speed of I billion items a second, even after billions of billions of years (and our earth has not existed for more than 10 billion years) not even one billionth of the total number of variants would have been examined.
But somehow an effectively functioning nerve net does form! And higher animals have not hundreds or thousands but millions of receptors and effectors. The answer to the riddle is concealed in the hierarchical structure of the nervous system. Here again we must make an excursion into the area of general cybernetic concepts. We shall call the fourth stage of evolution the stage of the complex reflex, but we shall not be able to define this concept until we have familiarized ourselves with certain facts about hierarchically organized nerve nets.
[1] I am generally following the report by S. E. Schnoll entitled "The Essence of Life. Invariance in the General Direction of Biological Evolution,'' in Materialy seminara "Dialektika i sovremennoe estestvoznanie" (Materials of the '"Dialectics and Modern Natural Science'' Seminar), Dubna, 1967.